
ICMSワークショップ
サステナブルな水産利用に向けた数理科学連携の可能性
気候変動や乱獲によって漁獲できる水産資源は減少傾向にあり,
日時と方法
- 2021年12月17日 13:00-16:00
- Zoomによるオンライン.参加登録フォームに参加登録をなさった方にのみ,当日の朝に具体的な情報をお知らせします.
プログラム
13:00-13:10 挨拶・主旨説明
13:10-13:50 松野孝平(北海道大学大学院水産科学研究院)
北極海における気候変動とプランクトン
13:50-14:30 壁谷尚樹(東京海洋大学 学術研究院 海洋生物資源学部門)
海洋生物における長鎖多価不飽和脂肪酸生合成酵素の多様性
14:30-15:10 小南友里(東京大学大学院農学生命科学研究科)
かまぼこの独特な食感を生み出すタンパク質メカニズムに関する研
15:10-15:20 休憩
15:20-15:50 パネルディスカッション(司会:齊藤宣一)
討論参加者
- 講演者3名
- 田畑公次(北海道大学電子科学研究所)
- 儀保 伸吾(理化学研究所数理創造プログラム (iTHEMS))
- 下川朝有(東京理科大学理学部第二部数学科)
15:50 まとめ・挨拶
※講演時間には、質疑応答、交代を含んでいます。
主催
東京大学大学院数理科学研究科 附属数理科学連携基盤センター
共催
九州大学マス・フォア・インダストリ研究所(文部科学省委託事業 AIMaP 受託機関)
世話人
- 齊藤宣一(東京大学大学院数理科学研究科)norikazu[AT]g.ecc.u-tokyo.ac.jp ([AT]は@に置き換えてください)
- 小南友里(東京大学大学院農学生命科学研究科)
- 時弘哲治(東京大学大学院数理科学研究科)
パネルディスカッションの記録
齊藤 これからパネルディスカッションを行います。司会はこのまま私、齊藤宣一(東京大学大学院数理科学研究科)が続けます。御講演者の松野孝平先生(北海道大学大学院水産科学研究院)、壁谷尚樹先生(東京海洋大学学術研究院)、小南友里先生(東京大学大学院農学生命科学研究科)には引き続きご参加頂き、さらにここからは、新たに、田畑公次先生(北海道大学電子科学研究所)、儀保伸吾先生(理化学研究所数理創造プログラム (iTHEMS))、下川朝有先生(東京理科大学理学部)の3人の先生方にも加わっていただきます。まずは、パネルディスカッションから参加の3人の先生方に、前半のご講演に対する感想などを伺えると幸いです。順番で田畑先生、いかがでしょうか。
田畑 まず自己紹介をしたほうがいいですかね。北海道大学の田畑と申します。現在の所属は北海道大学電子科学研究所と、あと化学反応創成研究拠点で特任助教をしております。専門は機械学習で、今、強化学習の一分野である多腕バンディットというアルゴリズムの研究を主にしています。機械学習というと、ラベル付きの訓練データが与えられて、それをもとに未知データに対して正しくラベルを推定するようなモデルを作るというようなことがタスクがよく扱われますが、強化学習はデータを自分で調べに行って、そこから得られた報酬を最大化する。試行錯誤しながらいい行動を選択するように学習するというアルゴリズムの研究をしています。その応用として癌の診断の高速化や化学反応の触媒発見の効率的な探索などをやっています。
今回のパネルディスカッションに関しては、機械学習以外の分野はそれほど知識がないので、なかなか新しく聞くことが多かったのですが、皆さん、非常に分かりやすく発表していただいて、どれもおもしろく聞けました。ディスカッションの中で何か提案するかもしれないですが、あまり意味のある提案ができるかどうか分かりません。よろしくお願いいたします。
齊藤 では、儀保先生、お願いいたします。
儀保 私も自己紹介からお話しします。今は理化学研究所の数理創造プログラム(iTHEMS)という、数学の人も物理の人も生物の人も理論をやっている人が集まる部署の生物グループの一員のポスドクとして研究をしています。バックグラウンドを少しお話ししますと、私は学生のころは学部から博士までは数学や生物ではなく、九州大学の芸術工学部の音響設計学科というところで音響工学、音について研究をしていました。ドクターを取ったあとに理化学研究所に移りまして、望月理論生物学研究室というところで生物の理論の研究に関わることになりました。特に体内時計みたいな振動現象というのは、音も振動するし、体内時計も振動するみたいな形で、数理の目から見ると共通のツールが使えるということで今研究しています。
研究のスタンスとしては、ほかのお二方は結構インフォマティクスなところが得意だと思いますが、私はどちらかというとシンプルなモデルを使って分かりやすいモデルで理解しやすい形で解析する、あるいは時系列データから重要な反応を予測するということを主にやっています。今日のお三方の講演は、資料をもらったときより、講演を聞いたときのほうがいろいろなデータが出てきて、すごく刺激的で、いろいろなデータが出てきたら、シンプルモデルより、むしろインフォマティクスのほうがいいのかなと思ったりするのですが、シンプルモデルが役に立たないとは思っていなくて、シンプルモデルとインフォマティクスの方法を組み合わせることで、分かりやすく予測のできるモデルとか、そういうところで貢献できるかなと考えながら見ていました。どうもありがとうございます。
齊藤 それでは、下川先生、お願いします。
下川 はじめまして。東京理科大学の下川です。本日は貴重な機会をいただきありがとうございました。簡単に自己紹介をさせていただきますと、私は数学科に所属しており、専門は応用統計学と機械学習と言われる分野になります。研究では医療データや、医薬データの統計解析に携わることが多く、本日のご発表では初めて見るデータや手法ばかりで、すごく勉強になりました。
3人の先生方のお話を聞かせていただいて最初に感じたのは、予測に特化したモデルを作って利用したいというご意見が非常に多いのかなと思いました。そういったものに関しては、近年研究が盛んにおこなわれている機械学習のアプローチを使うことで、なにかしらのいい結果が得られるかもというのが一つ目の感想です。
一方、予測ではなく、小南先生のお話にもありました、なにかしら裏にある背景等を、データを利用して解釈可能なモデルとして表現する際に、いわゆる古典的な統計学の手法や数学の分野を利用できそうと感じた次第です。どうぞよろしくお願いいたします。
齊藤 今の自己紹介の中で田畑先生と儀保先生はあとで話したいことがある、とおっしゃっていたような気がします。もし、よければ先にご発言していただけないでしょうか?
田畑 例えば松野先生の発表にあったプランクトンの話で、カメラを使ったプランクトンの濃度の計測では、ネットを使った計測とは違って、リアルタイムでデータをとるということになるんですかね。
松野 説明させていただきます。実際、アメリカのスクリプス研究所などで研究の方法はかなり進んでおりまして、実際にカメラを海の中に入れて、ハイスピードでフラッシュをたきながら水中ををジグザグに撮影していきます。カメラはケーブルで船上と繋がっており、リアルタイムに自動でどのプランクトンも分類して、数も数えることができますが、なかなか高額です。日本でもJAMSTECがやっているのですが、それを比較的簡単にといいますか、プランクトンを研究する人間は大学の教員から水産試験場などいろいろな方がおられまして、基本的に今でも顕微鏡を使っているのですが、そういう方でも簡単にできるようなツールがあればいいなと思います。例えば採集したものをシャーレに入れて写真を撮って、その写真の中の画像が自動で切り出される。そして、切り出されたものに対して事前に与えてある教師データを使い、自動でダーッと分けることができると、すごく便利だなと思います。ニュージーランドとかでもフリーのソフトで作ったりするというのを聞いたことがあるのですが、そういうこともできるのではないかと今考えているというレべルです。
田畑 ありがとうございます。僕はベイズ最適化を使った研究を行っているのですが、これは逐次的に計測を行うみたいな話で、ある点を計測して、その結果をもとにして、また次の別の点を計測するというような感じです。今回、実現可能性よりも新しいことと伺っていたので無責任に提案するのですが、例えばカメラと自律的に動くドローンみたいなものを組み合わせて、ある点を計測して、そこの計測結果をもとに、次にどの深さがいいとかどの場所がいいとか、自分で動いて計測するとか、そういうことが自動でもしできたとして、うれしいかどうか分からないのですが、プランクトンの濃度が場所によって違ったりする境界で何が起こっているのかというのを自分で調べに行くようなシステムが作れないかなということを、お話を聞いて考えていました。ちょっと意味があるかは分からないですけれども。
松野 それは実は非常に興味があります。一番最後のスライドで私が話していた、カメラを使って研究するメリットは、今まではネットを100メートル曳いて一つのデータ、つまり0-100 mで一つの絵でしかなかったものが、カメラであれば空間的に高解像度になるという点です。実際の海の中ですと、躍層と呼ばれる密度が急激に変わるような場所があって、そこにプランクトンがたまっている。物理や化学のセンサーではそういうものが見えるのですが、そこに何がたまっているかとか、どういうふうにたまっているかというのも分かっていないです。それを明らかにするため、カメラで撮影した動画を解析することで可能になると思います。しかもカメラを船から下ろすのではなく、自走するAUVに乗せてしまい、AUVが好きなところに行って、それが取ってきたデータがそのまま全部解析できれば素晴らしいと思います。ただ、人がマニュアルで解析することができない量のデータになると思いますので、そこで機械学習を使うというのはすごく有効だという印象を持っています。ありがとうございます。
田畑 ありがとうございます。
齊藤 儀保先生は、いかがでしょうか?
儀保 私はいつも時系列データを扱っているので、松野先生のご講演はいろいろな時系列が見られて、すごくおもしろいなと思っています。私がよくやるのは、単純かもしれないですが、FFTみたいなことをやって、プランクトンの変動にどういう周期が隠れているのかとか、そういうことを見るだけでも、周期と気候変動との関係性を見られておもしろいのかなと思いました。もう一つ、自分自身はそんなに明るくない分野ですが、予測という意味では、インフォマティクスの研究者が気象予報などの分野で使っているデータ同化という手法とLotka-Volterraモデルのような食う、食われるの相互作用のネットワークを使うことで何か予測可能で理解がしやすいものが得られるのかとも思いました。あと講演の最後でプランクトンの生活史も見ているとか、今後はデータ数を増やして解析していくとか、そういうようなところから、いろいろなデータから生物の個体数の増えたり減ったりの相互作用のダイナミクスのネットワークを捉えられたら面白いと妄想しながら聞いていました。
松野 私はまだ時系列データの解析自体がうまくできていないと自分で感じていまして、具体的に今お話しいただいたことはすごくやってみたいなと思いました。例えば最初におっしゃった周期性のところですが、季節が移り替わることで、プランクトンも増えて減ってというのは必ずあって、その周期性は捉えることができます。ただ、やはりデータ数がかなり必要といいますか、センサーではなく、プランクトンの種類まで分析する今の方法は人の手で行っているので、時間的に高頻度といっても1週間に1回と粗いのが実情です。それでも、イレギュラーなデータは外したりできるので、まずはデータをためていこうと考えています。世界に目を向ければ、昔から数十年単位でずっとデータをとっている地点もあり、データがオープンになっています。そのようなデータと、自身で得たデータを比較するということも可能になっておりますので、チャレンジするときに、ぜひ試してみたいと思います。またもしかしたら直接ご相談させていただきたいと思いますが、その時は宜しくお願い致します。
気象データについてですが、データ自体は日本全国どこでも非常に高解像度のものがあります。大気と海の中の生物との関係は、直接的ではなく、いくつもの物理化学過程を経た上での間接的な関係ですので、中々比較が難しいのですが、関係性を見いだせれば予測することができますので、検討していきたいと考えています。また、海のデータも今はオープンとなっており、使えるものがあるので、データの取り方を統一することで、自身のデータと比較することも考えていこうかなと思っています。ありがとうございます。
齊藤 それでは、どなたでも結構です。ご自由にご発言をお願いいたします。
下川 松野先生によろしいですか。最後にお見せいただいたGDMによる分析ですが、これは生物の種の多様性を幾つかの地点で観測してモデル化するということをしているかと思います。たぶん種の多様性の違いをモデル化したいということだと思いますが、モデル内に地形的な距離の影響は入れていないのではないかと思いました。資料を見せていただいていると、温度とか氷の解けている量とか、そういったものが情報として入れられていると思ったのですが、もし可能ならば距離の影響もモデル内に加える方が、よりいいモデルができるのかなと思ったのが1点です。
あと衛星データも使われていますが、これは自分が調べている領域内の全ての値が手に入っているが、実際にこのGDMによるモデル化をされているときは、おそらく自分が観測した地点の情報しか使われていない形だったと思います。そうすると、モデル化する際に、その他の地点における衛星データの情報を捨ててしまっているのだと思います。得られたモデルを適用して予測をする際には、全ての地点に対してその値を使っていると思いますが、モデル化する際には、捨ててしまっているのは勿体ないのではと思いました。例えば機械学習の分野でいったら半教師あり学習みたいな、応答のラベルはないのですが、特徴の値の分布の情報はあるというのを利用することで、よりよいモデルができるのではないかと感じました。
松野 距離のデータは入っているはずです。地点ごとのプランクトンの量を入れているはずなので、入っていると思います。
下川 物理的な距離の情報が入っているのかな?と思いまして。
松野 それは入っているはずです。二つ目の点はは、私の理解が追い付いていないです。面的には海域全体の表面のデータが衛星のほうはあるのですが、おっしゃるとおり、生物のデータはある地点でしかなくて、生物データがある地点に絞って、衛星データを抽出しています。
下川 特徴の値で、その地点の温度とかそういった情報をたぶん使われているのだと思います。せっかく他の地点の特徴値Xの情報があるので、応答Yはなくても、それをうまいこと使うという方法は広く行われており、そういったものを使えば、よりよいモデル化ができるのではないかと思った次第です。
松野 そんなことができるんですね。不勉強で知りませんでした。半教師あり学習というんですね。調べてやってみます。
下川 あともう1点、先ほどの時系列モデルのところも季節効果があるといったので、いわゆる古典的なモデルですが、シーズナルエフェクトを入れた形の時系列モデルを利用すれば、よりよいものができるのではないかと思いました。
松野 なるほど。時系列の解析については、実はモデルというところまでたどり着いていないです。データ数が足りないので、それは蓄えていって試してみたいと思います。分かりました。ありがとうございます。
齊藤 GDMによるモデルというのは、具体的にどうやって表現されるようなものなのでしょう。例えばモデルというと、われわれは方程式や公式、いろいろ表現のしかたがあると思いますが。
松野 方程式と言えばいいんでしょうか。下川先生がおっしゃったように、応答変数側があるプランクトンの多様度を計算しています。いろいろな生物の群集の多様度を計算していまして、それに対して、その現場での説明変数としては、水温とか衛星からのデータの氷の密度とか複数のパラメーターを入れて、一度モデルを動かしています。それが最初にあって、そこから生物データと関係のある説明変数だけ選ばれてきて、その説明変数の中の衛星データを使って予測を書くという流れになっています。
齊藤 その場合のモデルというのは、中身がブラックボックスになっているものなのか、一つ一つの効果に意味付けがなされているようなものなのか。
松野 意味付けはなされています。下図の横軸のPredicted Ecological Distanceというものが、ある地点とある地点の環境の差を数値化したもので、大きければその差が大きいというものです。先ほど話した説明変数の複数のパラメーターを足し合わせたものを横軸に取っているというようなイメージを私はもっています。
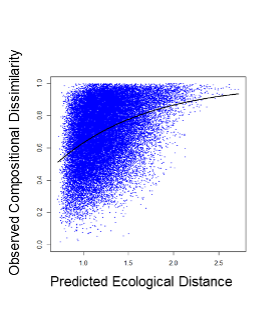
下川 少しだけ補足させていただきますと、通常の回帰と一緒で、今横軸で見せていただいているのは、いわゆる線形予測値をなにかしらの関数で変換して、一つの値にしたものが表示されている形です。通常の回帰とは応答のほうが少し特殊で、このモデルの場合は、サンプル2点間の違いを距離の形でモデル化していて、その違いを応答に持ってくるという形をしています。
齊藤 ありがとうございます。引き続き、何かご発言がございましたらお願いいたします。
儀保 先ほどのGDMについてもう少しお聞きします。あまり詳しくないのですが、線形モデルで回帰分析的なことをやるということは、重み係数を見ることで、重要な環境要因とかそういうものを推定できたりするんですか。
松野 選び出すことはできます。
儀保 それはこれからですか。
松野 いえ、お見せした結果は、既に重要な環境要因を選び出した後のものです。モデルを動かく際に、最初はより多くの衛星データを変数として入れています。ここでは氷の密度とか水温とか年間の一次生産量だけですが、ほかにも複数データを最初に入れておいて、そこから重要なものだけ、残ったものだけを使って予測をしています。ですので、その重み付けはできています。
儀保 重み付けをして、特にこれが必要だと特定したうえで予測に使う。
松野 そういうことをここではやっています。全部Rの中で動かしています。
齊藤 壁谷先生や小南先生のご講演や、あるいは壁谷先生、小南先生ご自身からのご発言でも結構ですが、何かございませんか。
小南 私の研究ではなく壁谷先生のご講演内容に関してですが、壁谷先生のご講演の中で、代謝ネットワークの形が違うことで代謝産物のプロファイルも変わるというお話があったと思います。注目している代謝ネットワークが、自分たちが想定している形どおりなのかどうかというのは、それこそ望月先生のモデルを使って検証できるのではないかと思うのですが、儀保先生いかがでしょうか。
儀保 私が岡田さんや望月さんの研究について自分が深く理解しているわけではないのですが、複雑なネットワークの中からあるシンプルなネットワークを取り出してくるというのが岡田さんや望月さんの研究だったと思います。確かにそういう意味ではもともと複雑だったものが淡水にうつった、海水にうつったで、重要なネットワークだけを残してシンプルな形に進化していくとかそういうことがあってもいいのかなと思ったりもします。
ただ、この代謝パスの場合は、ほかに枝葉があるのかがよく分からなくて。リニアにAがBになって、BがCになって、CがDになっているということで、そこで1個パスを止めると、そこだけが単純にただ途切れているだけに見えるので、岡田さんや望月さんがやっているような、複雑なネットワークから重要な部分だけを取り出すということにはなっていないように見えます。スライドではリニアにパスを書いているけれども、もうちょっといろいろループがあったり、いろいろなネットワークの中でこうなっているとかだったら、岡田さん、望月さんの話とも通じてくるのかなという気がします。
壁谷 今回のプレゼンではかなり省略した経路をお見せしているのですが、全ての経路を示すとかなり枝分かれのある形になっています。例えば最初の基質が不飽和化されるか、あるいは鎖長延長されるかで二つの経路があったりします。また、EPAからDHAの生合成に関しても、DPAを介して直接DHAへと合成される経路があったり、DPAからさらに鎖長延長されてTPAが合成され、それが不飽和化されたのち、β酸化すなわち鎖長の短縮を受ける形でDHAが合成される経路があったりするので、生物種ごとに非常に多様化していますね。
どのくらい多様化しているかというと、オレイン酸等の単価不飽和脂肪酸から、多価不飽和脂肪酸を生合成したり、オメガ6系の多価不飽和脂肪酸をオメガ3系に合成する経路は脊椎動物には一切存在しなくて、植物とか原生生物、一部の無脊椎動物だけに存在する毛色になります。脊椎動物の中でも、一つの遺伝子が複数の経路を担っている形になっていて、先ほど説明した経路の中でも、Δ6不飽和化とΔ8不飽和化経路は同じ遺伝子にコードされる酵素が触媒しているし、鎖長延長に至っては、炭素鎖が18と20ならこの経路上の全ての脂肪酸が一つの酵素の基質になり得ます。だから、一個の遺伝子が発現すると、同時に複数の経路が進行するという状態になります。ただ、それぞれの酵素に微妙に基質特異性があるので、同じ量の酵素が存在していた場合、ある経路を触媒する活性のほうが別の経路を触媒する活性よりも高いとか、そういう感じになっています。ですので、たぶんこういう遺伝子を何か別のもっと単純な酵母とか細菌で発現させて、全経路の一部分だけを取り出したような形で実験をすれば、時系列での実験も含めてデータとしてはたぶんとれると思います。
例えば複数の酵素を同時に酵母で発現させて基質としてある脂肪酸を加えると、それがインプットになるので、どんな脂肪酸がアウトプットとしてできてくるか時系列でモニタリングすれば、実験データとしてはなにかしら取れるとは思います。そこにモデルが適用できるかはちょっと分からないですね。結局、どちらにせよ、どこかにたどり着かなければいけない経路になっているというのがたぶん大事なのかなと考えています。ただ、活性を見ると、例えばこの酵素は炭素鎖を18から20へと延長するのですが、実はこの20が22までいってしまいます。だから、この図に書いてないところまで延長してしまっていて、その鎖長延長された脂肪酸はどこへ行ってしまうのかが分からないという状態です。もっと言うと、元の鎖長に戻るというパターンももしかしたらあるかもしれないという話で、その辺りは少し複雑かなとは思います。
儀保 すごくおもしろい情報です。確かにこういう複雑なパターンがあって、先ほどおっしゃったような酵素活性を変えたときの時系列も取れるということなので、先ほど言った岡田さんや望月さんの理論を使えば、ネットワークが分かっていれば、酵素活性を変えたときにネットワークのどの範囲までに影響を与えるかというのもネットワークから数学的に記述することができるというふうに私は理解しています。
例えば元々はいろいろなパスがあって、淡水や海水に適応する進化の過程で不必要なパスが落ち、重要なパスだけが残ったということが分かるかもしれません。あるいは、酵素活性を変える実験をしたときの時系列データが、理論的に影響が及ぶ範囲とは違うところまで影響を与えたとしたら、何か見つかっていないパスがあるのではないかということも推定できます。こういうフルのネットワークで酵素活性を変えた時系列を見るのは、岡田さんや望月さんの理論を使えば、おもしろいことができるかもしれないなと話を聞いて思いました。
壁谷 ありがとうございます。
齊藤 話はつきませんが、時間になりましたので、パネルディスカッションは、ここで打ち切らせていただきます。3人のご講演者の皆さまと討論参加者の3人の先生方、どうもありがとうございました。それから聴衆の皆さんも参加してくださいまして、どうもありがとうございました。このワークショップはここで終わりたいと思います。
以上